2021.10.01
児玉龍彦さんによると Covid-19 のウイルスは複製ミスが多いので、増殖速度が大きい変異種は自壊する、という。https://www.ric.u-tokyo.ac.jp/topics/2020/ig-20210824.pdf ここで根拠とされている理論が Manfred Eigen の提唱した Error catastrophe である。Eigen M. Selforganization of matter and the revolution of biological macromolecules, Naturwissenshaften, 1971;58:465-523. この論文は無料公開されていない(Abstract も無い)ので、大学の図書館にでも行かなくては入手できないが、Power Point の説明資料が見つかった。http://dosequis.colorado.edu/Courses/MethodsLogic/Docs/Eigen.pdf。更に理論を展開した論文や Review も見つかった。https://www.researchgate.net/publication/247004214_On_Eigen's_Theory_of_Self-Organization_of_Matter_and_the_Evolution_of_Biological_Macromolecules。後でじっくり読んでみようと思う。
・・・KAVLI INSTITUTE FOR THEORETICAL PHYSICS での Eigen の紹介はよくまとまっている。https://online.kitp.ucsb.edu/online/colloq/eigen1/:Manfred Eigen received his PhD at the University of Gottingen and is the former director of the Max Planck Institute for Biophysical Chemistry in Gottingen. In 1967, Eigen was awarded, along with Norrish and Porter, the Nobel Prize in Chemistry. They were distinguished for their studies of extremely fast chemical reactions induced in response to very short pulses of energy. For the last forty years Eigen has focused on the self-organization of matter and the evolution of biological macromolecules. His name is linked with the theory of the chemical hypercycle, the cyclicinkage of reaction cycles as an explanation for the self organization of prebiotic systems, which he described with Schuster in 1979. In 1992 he was awarded the Paul Ehrlich Prize for this work and its far-reaching consequences in biology. More recently his interest has shifted to the technological utilization of these ideas establishing a new "evolutionary biotechnology". Manfred Eigen is ranked as one of the leading internationally renowned scientists studying the molecular mechanisms of biological evolution.
・・・このサイトには、何と彼自身の講演のビデオがあった。まあ、論文を読まなくてもよいだろう。
・・・また、探したら、解説みたいな記事が見つかった。Commentary PNAS 13374-76 Oct.15,2002 "Error catastrophe and antiviral strategy" である。短いので最初に読むにはちょうど良いかもしれない。要約してみた。
・・・基本的な概念として、RNA や DNA に可能な配列全ての空間を考える。核酸は4種だから、長さを N とすれば、4^N の可能な配列がある。これが情報空間を形作る。これは物理的な時空間ではない。一つの軸には4つの点しかなくて、次元が N である。それぞれの点と点の間の距離は定義できる。いろいろ考えられるのだが、とりあえずは、核酸の種類が何個異なっているか、という風にも定義できる。一つの点(配列)は細胞という環境の中で複雑なプロセスを経て自らの複製を作ろうとするが、必ずしもうまくいくとは限らない。例えば、一つの配列 Sk と表記すると、この配列の i 番目の核酸がその通りに複製される確率 qik が定義できる。Sk の全体としての正しい複製の確率 Qk はこれらの積になるから、一つの核酸あたりの正常複製確率 qk がその幾何平均として定義できる。
Qk = qk^N = Π_i qik
である。qik は非常に1に近いと考えられるので、
Qk ≒ exp(-N(1-qk))
と近似できる。
・・・1つの配列 Sm があったとして、これが他の配列に比べて σm 倍複製速度が大きいとする。複製ミスも含めて、複製プロセスを表現するには、情報空間のベクトル Sk が Sk' に変換されるとすればよい。複製速度が大きいという事はその変換行列の対角成分が大きいということである。対角成分は σkQk であり、非対角成分は σk(1-qik) となる。ここで、k=m 以外の σk=1 としておけばよい。複製を繰り返せば、Sm の割合は Qm の比率で減っていくのであるが、他の配列よりも σm 倍複製速度が速いので、およその処 σmQm > 1 であれば、複製を繰り返しても維持できる。qm で表現すると、(1-qm) > ln(σm/N) ということになる。複製速度が遅いとか複製エラー確率が高いと、この条件を満たせなくなって、配列の優位性が保てなくなる。つまり自滅である。
・・・ウイルス対策としてのひとつのポイントは、複製速度を遅くするような薬ではなくて、複製エラーを大きくするような薬もあり得るということである。AIDS治療薬の開発においてこれはパラダイムシフトをもたらした、と最初の方に書いてある。1993年の論文である。
・・・単純化したモデル計算(Tarazona(1992)) が紹介されている(Fig.1)。この図はいろんなところで引用されているので、概念が判りやすいのであろう。核酸の種類は 2 とする。RNA の長さは 20 とする。全部で 2^20≒10^6 通りの配列がある。qik は全て同じで、一つの配列 Sm だけが他の配列よりも 10倍だけ複製速度が速いとする(σm=10)。1-qik に対して定常分布を計算している。ただし、この図では、Sm と比べて何個の核酸が異なるかという風に分類しているから、そもそも配列の数自身が二項係数になることに注意すると、1-qik = 0.11 を境にして、それ以上エラー確率が増えると配列の分布が完全に均一になってしまう。それ以下では、配列の分布には Sm を中心とする秩序が見らる。これは統計力学でいうところの秩序ー無秩序相転移に相当する。秩序パラメータとして consensus という量がプロットされているが、これは生物学的には配列が多少違っていても機能的に同じという意味である。この計算の場合は生物学的な対応が無いので、どう定義しているかは判らないが、おそらく、Sm に対して核酸変異の少ない配列を集めているものと思われる。
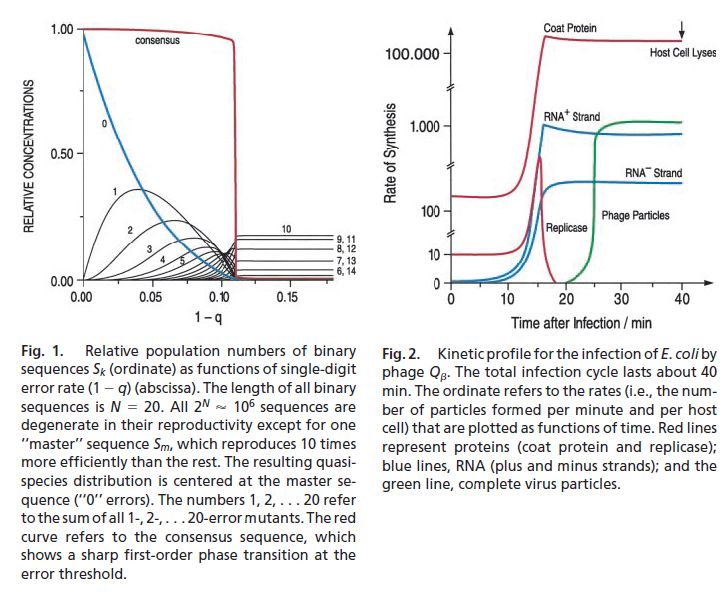
・・・以上の事は実は前置きであって、この Commentary の目的は現実のウイルス増殖との関係を考察することである。Eigen の論文としては、1991年に出ているらしいが、以下 M.Gebinoga という人の実験を紹介している。ファージというのは細菌を宿主とするウイルスで、実験がやりやすい。細菌にウイルスを入れて、1分置きに細菌を凍結して分析し、内部でのウイルス増殖の進行具合を測定している。Fig.2 である。ウイルスのRNAは m-RNA でもあるので、宿主細菌の装置によって読み取られて殻たんぱく質が作られるが、ウイルス自身の複製酵素が出来るまでに 10分位かかっていて、それが出来ると翻訳ー合成が本格的に進む。やがて、出来上がった殻たんぱく質が複製酵素のプログラム(遺伝子)に結合して酵素の合成が止まり、RNA全体と核たんぱく質が結合してウイルス粒子となり始める。後は一定の速度でウイルス粒子が増えていき、やがて細菌が融けて死亡する。大体10000-20000のウイルス粒子が生成し、それぞれが RNA と180個の核たんぱく質から成る。複製ミスがあるために、有効なウイルスは10%程度である。
・・・変異が促進される(エラー率が上がる)と優性配列の RNA が不安定化するが、野生型(wild type)はあまり影響されず安定である。
・・・中立的な変異もあれば、有害な変異もあり、また生存に有利な変異もある。これらは偶然である。
・・・Error catastrophe はあらゆる表現型の複雑な感染メカニズムに絡む。
Grande-Perez et al. の論文(2002)を引用している。『理論というものは複雑性を解決するものではない。理論は、どんな種類の規則的な振る舞いが期待できるか、また不規則性をうまく把握するためにどんな実験をすべきかを示すものである。』生物学においては物理学よりもこの教訓が大切である。Error catastrophe の理論はウイルスに対抗するために、増殖を阻害するのではなく変異を促進する、という新しいアプローチを提案している。このプロセスは免疫回避しようとするウイルスに挑戦するように見えるが、同時に病原性そのものの情報を失わせるかもしれない。個別のウイルス毎に多くの実験的研究が必要である。
・・・ということであるが、ここまでの話は増殖と変異の累積によって定常状態として適応した変異が固定化される、あるいは自滅してしまう、という話になっているので、児玉龍彦さんが言うように、少なくとも1ヵ月程度は盛んに増殖して他の変異を駆逐してから、自滅する、ということであれば、そういった変化はどうやって起きるのか、まさか途中で変異確率が上がるということでもないだろうから、依然として COVID-19 の流行波の盛衰の説明にはなっていない。それを示すには、定常状態ではなく、計算モデルでの時間経緯を見なくてはならないし、その経緯を実際の感染データと比較しなくてはならない。
<目次へ> <一つ前へ> <次へ>