


|

-目 次-
■SIRSとその問題点
■腸管出血性大腸菌(O157など)による溶血性尿毒症症候群の発症、進展に関する研究
■冬虫夏草:キノコと戦う昆虫たち -ヒトにも通じる昆虫の戦略-
■SIRSとその問題点
SIRS(全身性炎症反応症候群)は1991年にアメリカの胸部疾患学会と集中治療学会によって共同で定義された炎症反応の新しい概念である (Ref.1)。以下の4項目のうち2項目以上を満たす場合をSIRSとする。1)
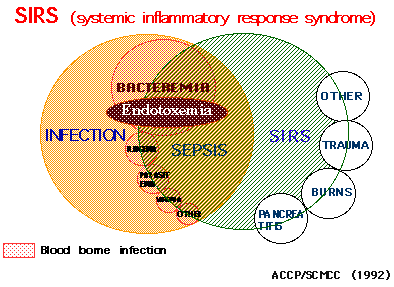
体温 >38℃ or <36℃、 2)脈拍 >90/min、3)呼吸数 20/min または PaCO2<32 torr、4) 白血球 >12,000/mm3 または <4,000/mm3または>10% 未熟型(band)。SIRSは図1にあるように外傷、熱傷、膵炎、侵襲の強い術後など感染を伴わない全身性炎症と、感染を伴う全身性炎症を包含する。そして感
染を伴うSIRSがsepsisである。これ以前にもBoneが菌血症が見られなくともどこかに感染があり全身性炎症反応が認められるときを sepsis
syndromeを定義している(Ref.2)。
従来、敗血症は菌が血中に侵入し、重篤な全身症状を呈する状態であると定義されてきたが、わが国では、早期からの抗生物質投与の影響もあって、一般的に血液培養の陽性率が低く、敗血症と確診することは困難な場合があり、多くは敗血症疑いと診断される。従ってsepsis syndromeは敗血症といわゆる敗血症疑いを含む概念といえる。さらに、SIRSではsepsis syndromeの定義をベースにして、感染によらない炎症反応を加えてより緩やかな診断基準を設定している。従って、SIRSと診断された患者は多彩な病気の集団であるといえる。
SIRSは炎症学の立場でみると SIRSは高サイトカイン血症として理解されよう。SIRSでは血中の炎症性サイトカインは高値(hypercytokinemia)を示す。しかし、感染を伴うと一般に血中サイトカイン濃度はより高値を示す。感染症学の立場で見た場合、SIRSは安易な定義とも思える。SIRSは侵襲による全身炎症反応をしめす患者の臨床治験のentry
criteriaとして提唱されたようである。
SIRSには感染を伴わない場合、感染を伴うが菌血症を伴わない場合(局所感染)、菌血症を伴う場合の3つの型があると考えていいい。後2者はいずれも
sepsisと定義されるが、菌血症があるか否かは重大である。細菌が多臓器に播種されることを考えなければならないから、血液培養はこれまで通り施行すべきである。
エンドトキシンに関して言えば、我々の成績ではエンドトキシンに特異的な方法で陽性の真のエンドトキシン血症はグラム陰性菌による感染症でのみ認められる。SIRSでは図に示すように感染を
伴うSIRSの一部でのみ認められる(Ref.3,4)。もし、エンドトキシンに非特異的な方法で測定する成績をこの図にあてはめるとエンドトキシンは非常に広範囲なSIRSで認められることになる。
図 SIRSにおけるエンドトキシン血症の位置(稲田)
References
1. Members of the American College of Chest Physicians/Society of Critical
Care Medicine Consensus Conference Committee, Difinitions for sepsis and
organ failure and guidelines for the use of innovative therapies in sepsis.
Chest 10:1644-55/Crit Care Med 20:864-74,1992
2. Bone RC, Crit Care Med 17:389-393,1989
3. 遠藤重厚、稲田捷也、エンドトキシンと病態, へるす出版, 東京, 1995.
4. 稲田捷也、遠藤重厚、SIRSにおけるエンドトキシンや外来抗原に対する反応性の変化 -SIRSの動物モデル確立の必要性- エンドトキシン研究1
基礎と臨床、菜根出版、東京, pp97-103,1998.
■腸管出血性大腸菌(O157など)による溶血性尿毒症症候群の発症、進展に関する研究
血清型O157などの腸管出血性大腸菌による溶血性尿毒症症候群(HUS)は死亡率が高い。白血球増多が危険因子とされてきたがそれの病因への関与につい ては殆ど考慮されてこなかった。重症HUS患者ではサイトカインのうち好中球遊走活性化を担うインターロイキン8(IL-8)と、好中球活性化に伴い放出 される顆粒球エラスターゼが極めて高値であることがわかった。顆粒球エラスターゼは抗菌性を持つ一方で組織障害因子でもある。ベロ毒素による障害された 腎に好中球が集積し、ここで好中球が二次的に活性化され顆粒球エラスターゼ、活性酸素が大量に産生され腎障害を増悪させると考えれれる。
このことは、HUSは腎での障害が顕著にあらわれた多臓器不全の亜型であることが推察される。
著者らはベロ毒素が好中球を活性化することフローサイトメトリー法で明らかにした。HUSの発症にベロ毒素による好中球活性化が関与していることを示唆している。
■冬虫夏草:キノコと戦う昆虫たち -ヒトにも通じる昆虫の戦略-
ヤンマタケの子実体が生えたトンボや、長々と伸びた柄をもったキノコ(トビシマセミタケ)が土中のセミの幼虫から生えている異様な写真をキノコ図鑑でみた
とき、これは昆虫の真菌症であると合点する。昆虫類にいわゆる真菌症を起こすものは、殆どが子のう(嚢)菌亜門、バッカクキン科、冬虫夏草属Cordycepsに含まれる。真菌はヒトをはじめ多くの動物、植物に戦いを挑むが、昆虫も例外ではないようだ。これに対して昆虫はたとえ短い命でも必死になって敵に向かっていくに違いない。
脊椎動物が敵に立ち向かう術には免疫自然免疫と獲得免疫の2つがある。前者は相手に対して特異性がなく常時備わっている防御系であり多くの液性因子や貪食
細胞(単球・マクロファージや多形核白血球(好中球)など)が担っており、後者は特異的であり微生物が生体内に侵入した時に獲得する防御系である。獲得免
疫系の担当リンパ球であるT細胞やB細胞は、分化の途上で遺伝子再編成によって個々の微生物などがもつ抗原ペプチドなどに特異的な受容体を持つクローンを
形成する。微生物が侵入すると対応したクローンが認識しB細胞は抗体を産生し、T細胞は細胞障害性T細胞などになる。
自然免疫の真髄は、外来異物を認識して貪食・排除することにあるが、それを担っている貪食細胞がどのような認識機序で微生物を捕らえているかは長い間不明
であった。最近になって、これらの細胞が受容体を介して微生物表層の分子構造を認識することがわかってきた。その受容体はパターン認識受容体 pattern
recognition receptors(PRRs))と呼ばれ、微生物の共通性の高い繰り返し単位を認識する。この単位をpathogen-associated
molecular patterns; PAMPsと呼んでいる。PAMPsにはグラム陰性菌のリポ多糖(LPS、内毒素)、グラ陽性菌のペプチドグリカンやリポタイコ酸、そして真菌のβ-グル
カンなどがある。
PAMPsを認識するPRRsの主なものを挙げてみると、
●C型レクチン:糖との結合にCa2+を必要とする動物レクチンをC型レクチンと呼ぶが、そのなかで、マクロファージマンノース受容体やコレクチンなどはPRPsである。β-グルカンを認識するDectin-1も最近報告された。
●補体受容体(CR3):補体C3bの受容体であるが、β-グルカン、結核菌、百日咳菌、リューシユマニア、ヒストプラスマなどの微生物糖鎖を認識するPRPsである。
●CD14:リポアラビノマンナン、リポタイコ酸、マンノウロン酸重合体、ペプチドグリカン、大腸菌などと結合する。
●スカベンジャー受容体:マクロファージが持っており、本来の機能として酸化、アセチル化、糖化などの修飾を受けて変性したLDLを認識する。その結果こ
の細胞は泡沫化されるが、PRRsとしても働き、微生物のLPS、リポタイコ酸、大腸菌などとも結合する。
●トール様受容体(toll-like receptors;TLR):TLRは膜上に最も整然と準備された受容体レパートリーである。TLRには少なく
とも10種ある。TLR1はTLR2と強調し、トリアシル基を有する細菌由来のリポペプチドを認識する。TLR2はペプチドグリカンやリポペプチドを認識
する。TLR2欠損マウスは黄色ブドウ球菌の感染に対する感受性が高まる。二本鎖RNAはインターフェロンα・β産生の強力な刺激になるがTLR3を介し
ている。TLR4はLPSの受容体であり、長い間不明だったLps遺伝子はTlr4geneそのものである。LPS受容体はLPS・LBP複合体を認識す
るCD14とシグナル伝達系としてのTLR4から構成されていると考えられる。TLR5は細菌の鞭毛タンパク質フラジェリンを認識する。TRL6は TLR2と会合してリポペプチドを認識する。TLR7やTLR8は単鎖RNAと結合する。TLR9のリガンドは強力な免疫賦活作用を持つ微生物DNAのシ
トシンとグアニンの回文配列(CpG配列)である。
以上のように脊椎動物の貪食細胞は、PRRsを介して糖鎖をはじめとする微生物成分PAMPsを認識する。その結果、活性酸素や抗菌物質、さらにサイトカ
イン(主にTNF-α、IL-1β、IL-10、IL-8、GM-CSFなどの炎症性ないし炎症関連サイトカイン)とよばれる細胞活性化蛋白を産生・分泌
し、自己分泌、傍分泌によって貪食細胞自身を活性化し、自然免疫の成立に寄与する。
一方で膜表面だけでなく血液中にもPRRが存在する。中でも重要なのがC型レクチンに分類されるMBP(マンノース結合蛋白、マンナン結合レクチ ン:MBLともいう)である。MBPは補体のレクチン経路を始動させる働きがある。MBPは二つのセリンプロテアーゼ(MASP-1(MBL-
associated serineprotease-1)とMASP-2)の複合体であり、多くの細菌、酵母、ウイルスにある糖鎖の繰り返し構造と結合する。すると、補体のC4、C2が活性化され、C3からC9までのカスケード反応が次々と進行する。最終的に活性化補体成分が集合して膜侵襲複合体が作ら
れ、細菌表面に陥入し、細菌を死滅に導く。また活性化の途上で産生される白血球遊走因子は血管から白血球を感染巣に集積させる。同様にアナフィラトキシン
はマスト細胞に働いて脱顆粒してヒスタミンを放出させ、結果的にヒスタミンにより血管透過性亢進がもたらされ、白血球集積を助長する。活性化補体の一部で
あるC3bは微生物表層に結合し、微生物C3b複合体は白血球のC3b受容体に結合し効率よく取り込まれる。これによって白球球は活性化され炎症性サイト
カインが産生される。MBPの構造遺伝子の第1エクソンにおける3つの点突然変異をもつ小児では、血中MBP濃度が著しく低下し、重篤な感染を繰り返す。
補体の活性化にはこの他C3(H2O)と微生物抗原との結合による活性化経路(別経路)、抗原抗体複合体による古典的経路があり、互いに補完しあってい
る。
細胞膜上のTLRを始めとするPRRsと、補体を中心とするの液中の認識因子はどちらも重要であり、両者は互いに補完しあいながら働いていると考えられる
(TLRが最近の流行的研究になっているのは止むを得ないとしても、感染防御における補体の重要性が無視されがちなのを憂う)。
さて昆虫類はどのような免疫系をもちキノコに対処しているのだろうか?冬虫夏草の真実に迫ろう。
昆虫類の感染防御に関する断片的研究によれば、昆虫には特異免疫系はないが(免疫グロブリンの遺伝子は軟骨魚類から出現する)、自然免疫系が存在する。補 体のレクチン経路は、ホヤなどの脊索動物にも哺乳類とほとんど同じ遺伝子群が存在しているが、ショウジョウバエでも補体関連遺伝子が確認されており、昆虫 類は原始的な補体系をもっていると考えられる。先に述べた微生物成分を認識するレパートリーとしてのTLR群は、始めはショウジョウバエ(Drosophia melanogaster)の発生初期における体形成に関わっている蛋白tolllとして発見された。そしてこの遺伝子に変異を持つショウジョウバエが真菌感染に罹りやすいことがわかり、あっという間にヒトの感染防御まで研究が進んだ。
昆虫は、傷や菌の感染に伴い、TRLなどを介して微生物成分を認識し、体液中に抗菌・抗真菌タンパク(例えばセクロピンやアタシン)を分泌する。現在ま
で、ハエやガを中心に様々な抗菌タンパクが単離され、それらの抗菌タンパク遺伝子がゲノム中で大きなファミリーを形成していることも明らかになっている。
昆虫の血液が空気に触れると黒変するのは黒色色素(メラニン)が合成されるためで、酸素の存在下でチロシンやドーパなどのフェノール性物質が酸化され、そ
の結果できたキノン類が重合してメラニンができる。この反応を触媒するのがフェノール酸化酵素である。微生物のPAMPsを液中や膜上のPRRs蛋白が認識しても、フェノール酸化酵素系が活性化され、最終的にメラニンが微生物周囲で合成される。その過程で産生されるキニンは周囲の高分子物質をなめし効果で
強固にして微生物を閉じ込めたり、それ自身が抗菌的に働く。PRRsとしてペプチドグリカン認識蛋白(PGRP)、b1,3グルカン結合蛋白 (BGBP)、LPS結合蛋白(GNBP)などが報告されている。PGRPは昆虫からヒトまでその遺伝子の存在が確認され、実際昆虫でも蛋白が機能してい
る。BGBPはカイコやザリガニ、カブトガニ、ブラックタイガーエビなどに存在している。GNBPはLPSのほかグルカンにも結合するが、昆虫類全般に存
在する。これらの認識蛋白は貪食細胞膜上に存在し、PAMPsと結合すると脱顆粒、抗菌蛋白の合成が促される。
カイコの血液(SLP)中にはPGRPとBGBPが存在し、それぞれPGやβ1,3グルカンによってフェノール酸化酵素系が作動し、最終的にメラニンが形
成される。この現象を利用して、これらの微生物菌体成分の定量ができるようになった(SLPテスト)。また、カブトガニ血球(アメーボサイト)の抽出液中にはLPS結合蛋白(C因子)やβ1,3グルカン結合蛋白が存在し、LPSやβ1,3グルカンと結合するとカスケード反応が動いてゲル化がおこる。これを
応用して日本の試薬メーカーがLPSやβ1,3グルカンを個別に定量できるキットを開発し、グラム陰性菌敗血症や深在性(臓器)真菌症の診断に用いられる
ようになった。LPSは内毒素(エンドトキシン)とも呼ばれ、TLR4を介してマクロファージを活性化し、免疫能を亢進したり、血液凝固を促進したり、多
彩な生物活性を示すが一方で敗血症性ショックの原因とも考ええられている。筆者はエンドトキシンに特異的は測定法を用いて、敗血症を含む種々の病態におけ
るエンドトキシンの役割を研究し、エンドトキシンと病態における新しい概念を構築した。別の機会に述べたい。
ヒトゲノムは約30億個あるが,ショウジョウバエもヒトの約6%1億8000万個のゲノムしか持たないのに、ヒトにひけをとらない自然免疫系をもつことは驚異的である。
昆虫はキノコの攻撃に対して彼らのなしえるすべての手段で対抗し、最後はあえない最期を遂げるが、キノコは昆虫の栄養分の他に昆虫が感染を免れるために産
生した抗菌物質なども当然のように吸い上げていくに違いない。冬虫夏虫はその全体像なのである。
(2006年3月)

|
|
Copyright (C) Katsuya INADA 1997-2012, All rights reserved.
このサイトを利用することにより発生する損害等については一切責任を負わない
|
|