
伊藤 浩司
1.はじめに
北海道の植物についての知識やデータが豊富に集まるにつれ、北海道における植物の分布形態についても本格的な論議が必要になってきている。
北海道やその周辺地域の植物相の特色は、舘脇の一連の研究、例えば“北太平洋諸島におけるシダ類の分布”(1951)、“隔離分布の研究”(1954)、“北太平洋諸島のラン科植物の植物地理”(1954)、“北太平洋諸島のサクラソウ属の植物地理”(1955)によって明らかにされてきた。しかし、これらの研究は主に高山域や戦前未知の研究領域であったサハリンや千島あるいは交通の不便ないわゆる原生林域であった。しかし、最近交通網の発達によって道内至る所に歩を進めることができるようになり、低地や湿原からも新しい植物の報告が相次ぎ北海道の植物相についての再検討を必要とするような趨勢になってきている。
1953年大井次三郎の“日本植物誌”第1版出版以後の北海道新産植物の主なものをあげると次のようである。
湿原:フサスギナ(スギナ科)北見、大雪山、カラクサキンポウゲ(キンポウゲ科)霧多布、ハナタネツケバナ(アブラナ科)小清水、霧多布、カラフトネコノメソウ(ユキノシタ科)阿寒、霧多布、カラフトホシクサ(ホシクサ科)浮島
山地:タカネハナワラビ(ハナワラビ科)有珠山、ミヤウチソウ(アブラナ科)宗谷、旭川、ヒロハガマズミ(スイカズラ科)定山渓天狗岳、日高山地、カラフトイワスゲ(カヤツリグサ科)崕山
森林:ヒメホテイラン(ラン科)定山渓、カラフトキバナアツモリソウ(ラン科)礼文島、北見端野、ベニシュスラン(ラン科)苫小牧、ヒナチドリ(ラン科)日高、釧路白糠、ウラホロイチゲ(キンポウゲ科)十勝
高山域:スガワラビランジ(ナデシコ科)十勝ニペソツ、オオウサギギク(キク科)石狩クマネシリ、ピヤシリ山
北海道の植物の分布については舘脇(1960)、渡辺・大木(1960)の研究がある。しかし、北海道では未だ植物地理学の立場から論じられている研究は皆無といってよいであろう。これからの研究は論理と原理に寄って導かれたものでなければならないのである。
2.北海道植物の分布の形態
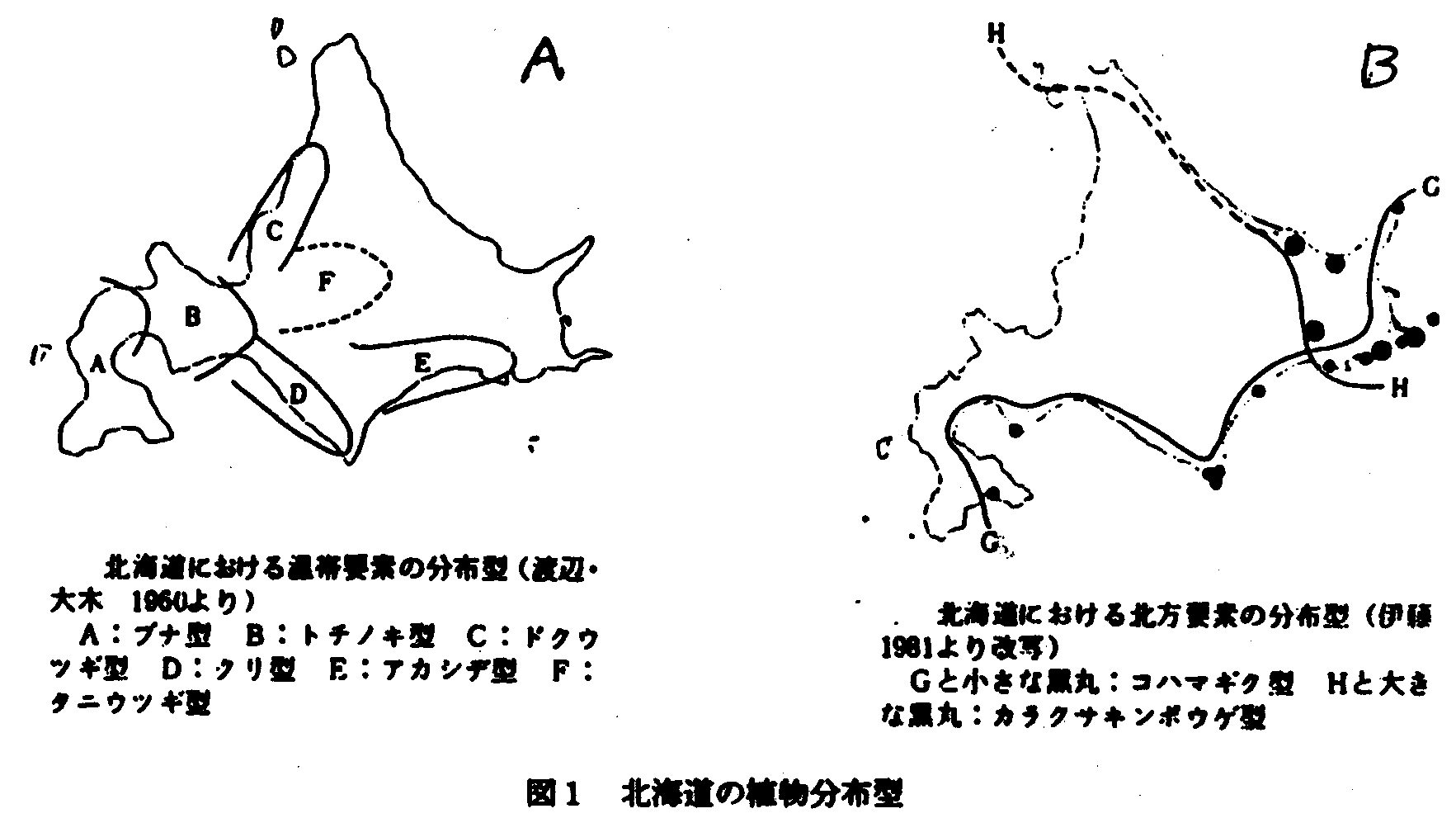
図1に示すように現在の植物の分布の形態を要約するひとつは北進型であり、今ひとつは南下型である。この北進と南下は第4紀における氷期と温暖期の交代に原因がある。現在は氷期と氷期との間の温暖期であり、南の温かい地域に生活の本拠を持つ植物が気温の上昇と土などの環境条件の変化によって、次第に北進しつつある状況下にある。これに反して、寒冷期に栄えた寒いところに生活の本拠を持つ植物は気候が次第にかれらの生理的要求にあわなくなり、生活を続けるころが出来なく、また南からの植物との競争に破れて消滅するが、特殊な場所に追い詰められて、ほそぼそと生存しているにすぎなくなる。したがって、現在の気候条件下で分布領域を広げつつある温帯性植物の分布の形態と北方の植物の低地における分布の形態とを同一にみることは出来ないのである。演者は前者に対しては分布型と呼ぶが、後者に対しては遺存パターンと呼ぼうと考えている。分布型の主対象となる植物と遺存パターンの主対象となる植物の違いについても多少の考察を必要とする。ラウンケアの生活型の統計結果から、ラウンケアは所謂生活形気候を提案したが、その中で熱帯地方は高木気候で占められ、ツンドラ地域は地表植物気候が優占する。このような違いはひとつの仮説として結果する。木(Trees)は草あるいは下床の植物よりも気候変化に一層敏感であり、草や下床の植物は気候変化に対しては比較的保守的である。このことは広く植物の分布型を論ずるには樹種を対象とすることを暗示する。
このような観点にたって、図1により北海道植物の分布をまとめてみると次のようになろう。
A. 北進分布
1.ブナ型 2.トチノキ型 3.ドクウツギ型 4.クリ型 5.アカシデ型 6.タニウツギ型 (渡辺・大木1954)
B.遺存パターン
1.カラクサキンポウゲ型 2.チシマコハマギク型 3.ミヤウチソウ型 (伊藤1983,1994)
C.不連続分布型
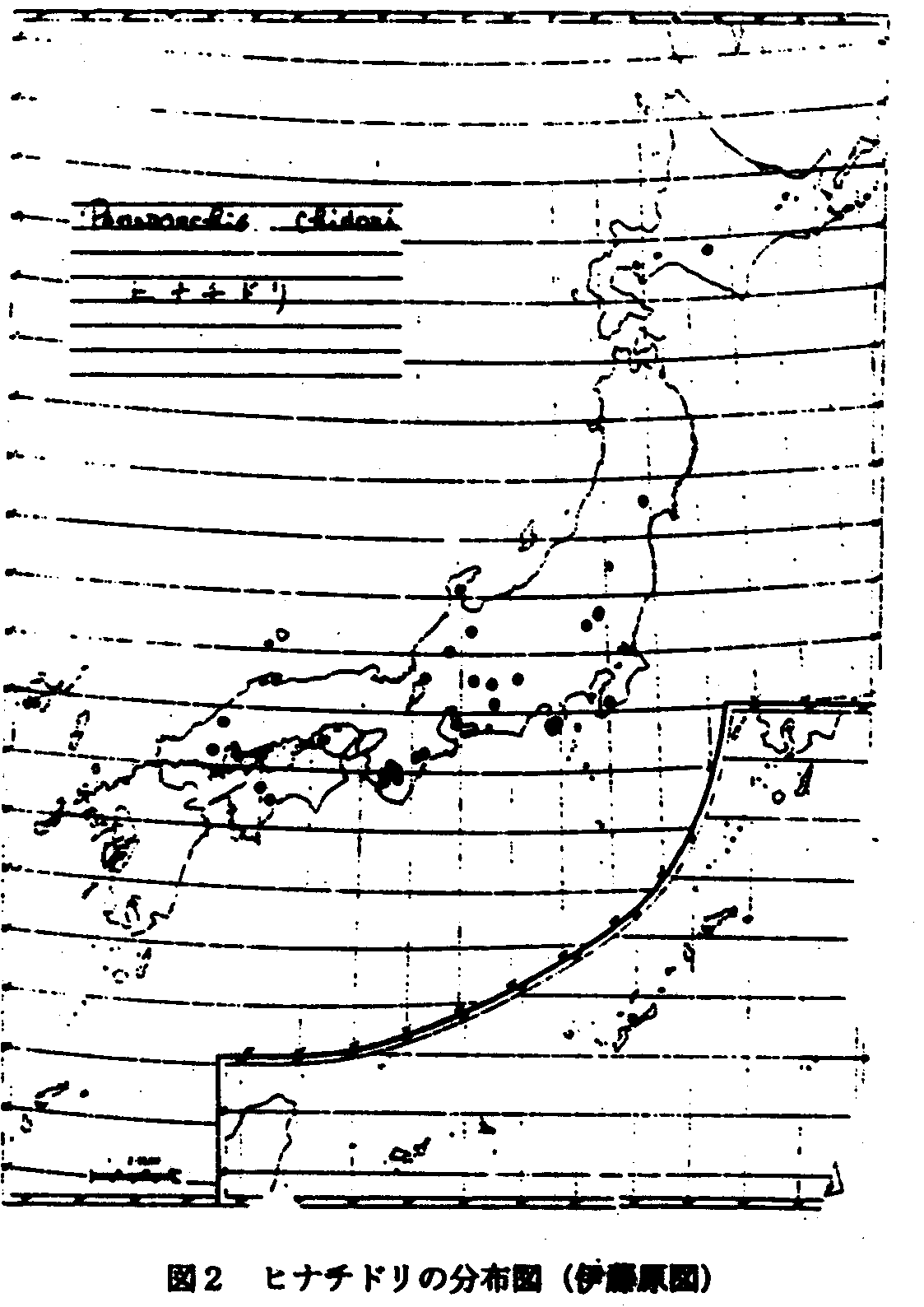
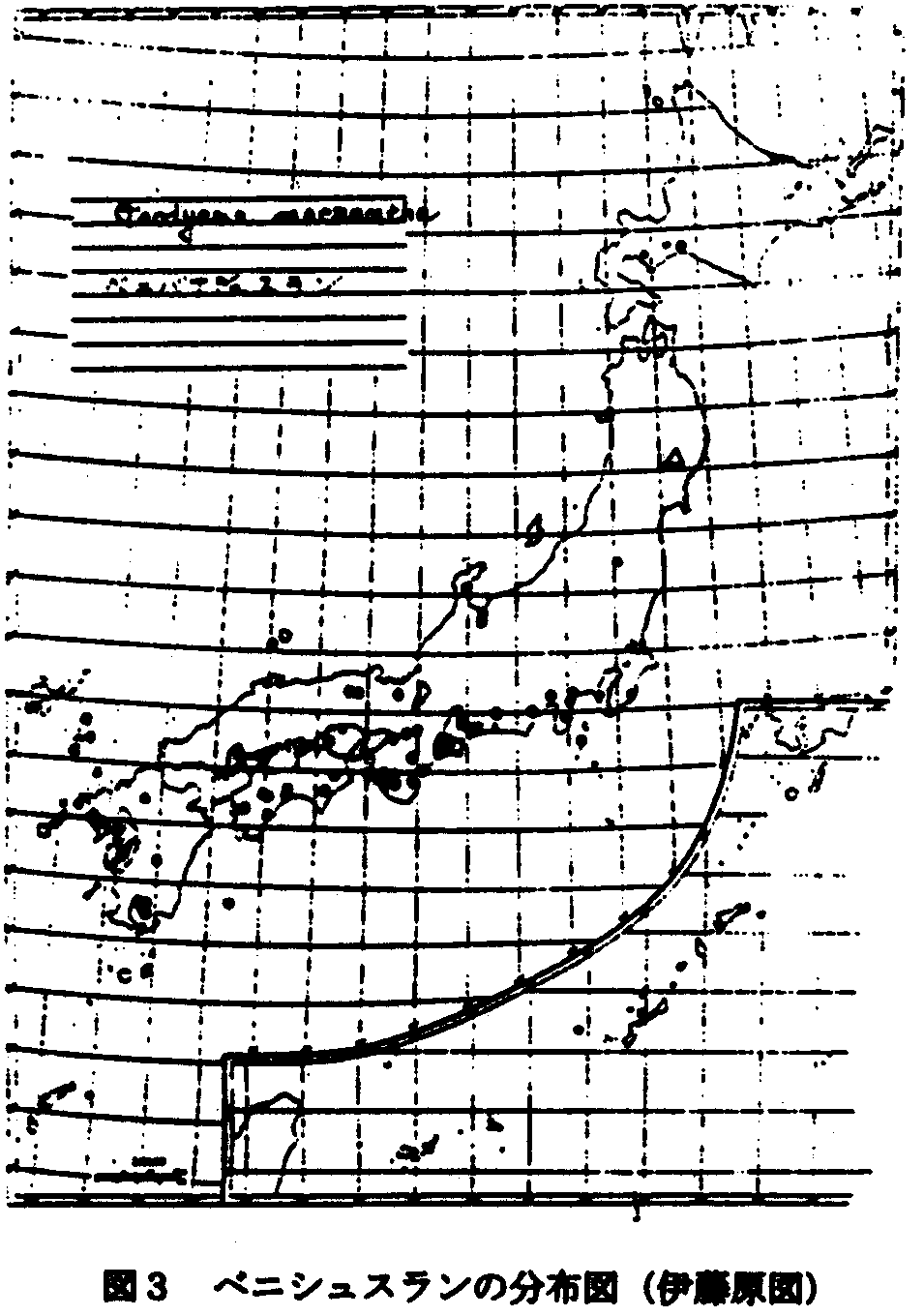
1.陸中ー日高型:ヒナチドリ(図2)、ベニシュスラン(図3)をはじめヒダカミツバツツジなどがあり、分布型からみればクリ型に含まれよう。 (舘脇1954)
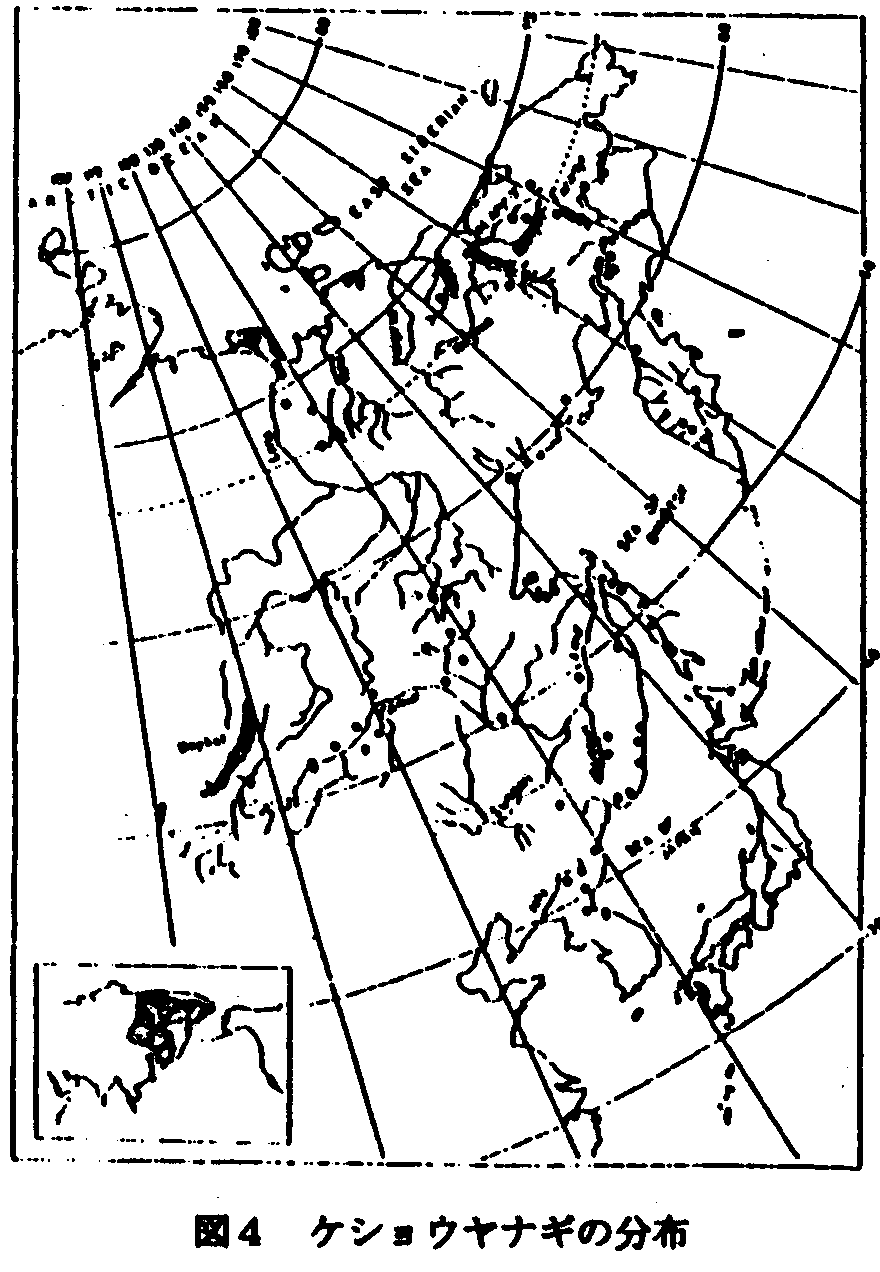
2.ケショウヤナギ型 (伊藤1994)
しかし、植物の分布形態の発現はいろいろ複雑な要素の絡み合いによってできているものであり、基本的には氷期以降の気温の変動やそれに伴う土壌の栄養状態や乾湿性の変化、さらに植物と生活をともにするような動物―食植動物、花粉媒介昆虫・小動物、土壌生物・微生物、種子運搬者など―の存在や活動を無視することはできないであろう。さらにまた、日本のように南北に山脈が走り、冬には大陸の乾燥空気を一気に湿らせる日本海の存在は冬季大量の雪と湿った空気、それによる植物体の保護、植物体への荷重圧と比較的温暖な気温に恵まれている日本海側とその反対の太平洋側とに植物の分布や分化に影響を与え、二次的な植物分布形態をつくりだすのである。上述のタニウツギ型などは多分にそのような影響下の分布型である。
3.興味ある遺存種の溜り場
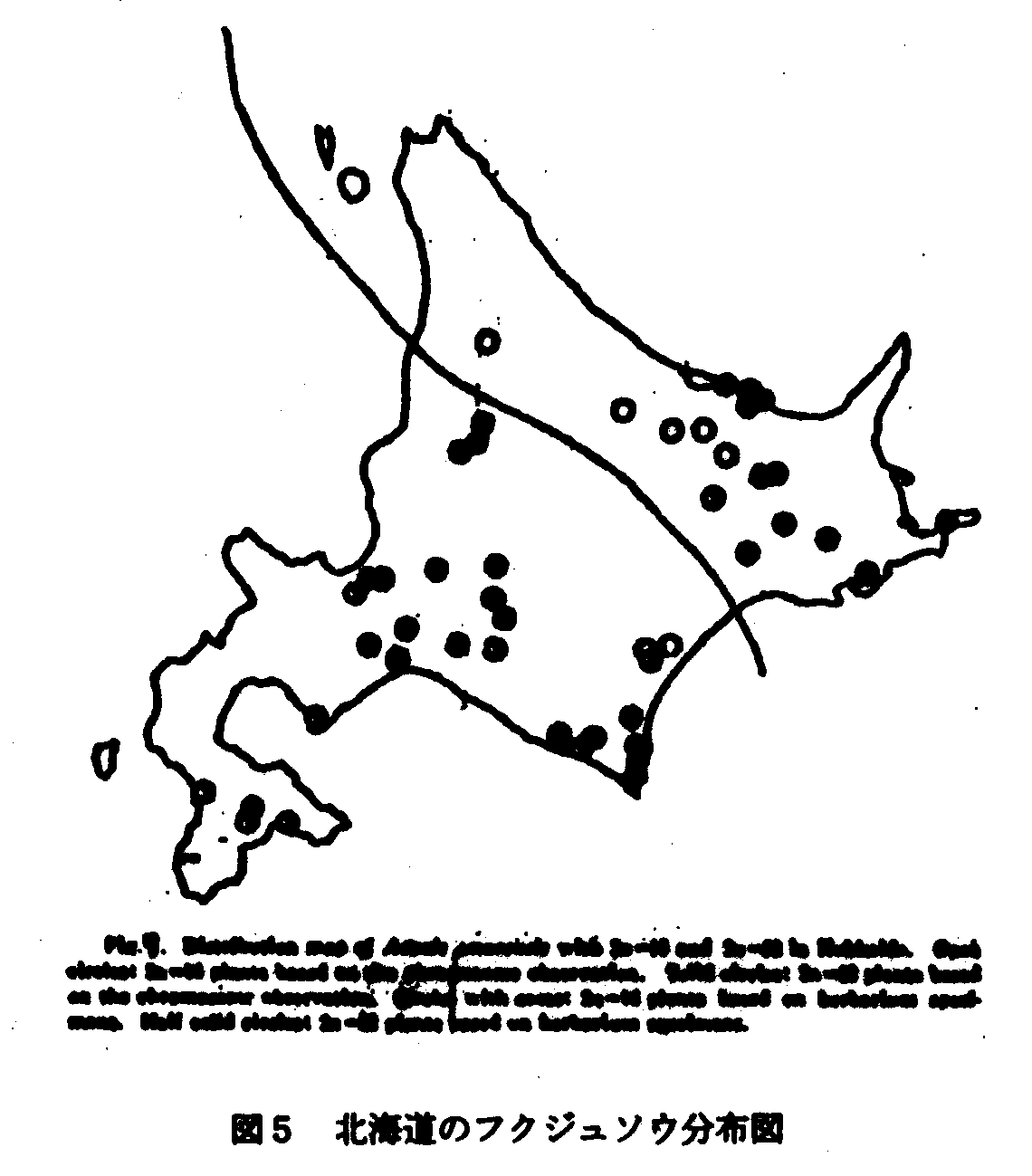
北海道の植物相は留萌ー襟裳岬を結ぶ線で2大別される(伊藤1988)が、個別的に植物分布の様子をみると、注目されるのは襟裳岬付近を中心とする日高南部とその周辺地域である(図4)。ここではアポイ山の北方性要素の蛇紋岩変性植物群に加えて、日高・十勝にまたがってケショウヤナギ、十勝側にはアムールイチゲ(ウラホロイチゲ)、少し北上して富良野付近にはエゾオオケマンのようなサハリンの植物と密接に関連する植物がみられるほか、ヒダカミツバツツジのような南の本州要素(陸中ー日高型分布の延長線上とも考えられるが)などサハリンからの植物と本州からの植物とが混在している地域である。花粉分析の上からあまりに大量のブナの花粉がでるので、この近傍にまだブナが生存しているのでないかとの疑いをもたれた地域の一部でもあると考えるとこの地域は北海道における植物の過去寒冷期から、現在の温暖期にいたるまでの植物の分布経路を考察する際に見逃してはならない地域であると推測している。寒冷期には襟裳岬の最先端まで北方性植物は南下してきたであろうし、高さの割にアポイ山における北方性植物の豊富なことをおもえば、その当時でも気候条件は極めて厳しいものであっただろう。しかしまた、日高山脈の切れるわずかな低地は温暖期にいたり南西北海道から北進してきた植物のひとつの重要な通路であり、アカシデ型分布を生みだす結果になったのである。草本種でもオオサクラソウとエゾオオサクラソウの分布、フクジュソウとエダウチフクジュソウの分布を考える際この通路は意味のある存在である。いわば、北海道の南端部は現在は温暖地の植物の分布の第一線であるとともに、なんらかの理由で過去温暖期の植物の遺存地帯であったかもしれず、また、同時に過去寒冷期の植物が必死になって生存の場を確保しつつある現状にある。したがって、これら遺存的北方性植物は低地にあっては気温の上昇とともに、豊富な温帯植物との激しい競争に打ち勝たねばならず、種の保存上難しい問題を我々に提示しているのである。
4.気候変動と植物分布への影響
気候変動は植物の移動や分布型の成立にどの様に作用しているであろうか。まだ荒ら削りの段階であるが一応の試論を示してみようと思う。(表1)
極めて仮説的であるが、現在の植物の分布の不連続性を考える場合には過去における地形的変動のみならず、土壌の有機物分解、腐植の形成、酸性度の変化などの環境条件の総合的考察が必要であり、図面上に分布の点を打つことだけで終始してはならないのである。
表1 気候変動と植物分布への影響
| 気候変動 | 南北植物の関係 | 地形・土壌的変化 | 遺存関係 |
|---|---|---|---|
| 温暖化 | S>n | 湿潤ー乾燥-中生 | 遺存的要素は湿潤型;断崖の縁 |
| 低地の湿潤地の消滅 | 岩地の縁、突端 | ||
| 新砂丘の形成ー湿原の発達 | |||
| 前植生は草本、矮性木本、開放性、土壌有機物の分解早急、土壌塩類の豊富化 | |||
| 樹木植生からの開始 | |||
| 寒冷化 | N>s | 乾燥ー湿潤ーツンドラ様凍結 | 遺存的要素は中生ー乾燥地 |
| 低地の露出、新砂浜・砂丘の形成ー湿原ーツンドラ断崖・海岸断崖 | (ブナの石灰岩地への逃避)温泉・火山地帯 | ||
| 前植生は木本・森林、閉鎖性、 | 温泉・火山地帯 | ||
| 土壌有機物分解は遅延、停止、 | |||
| 泥炭化、地表植物の増加、 | |||
| 植生推移の停止、 | |||
| 永久社会の形成 |